Parameter Estimation for Biochemical Reactions in Phototransduction
Vasilios Alexiades
Department of Mathematics, University of Tennessee Knoxville, TN 37996
and
Oak Ridge National Laboratory, Oak Ridge TN 37831, USA
Abstract
Vision begins when photons are captured by rhodopsin
molecules in photoreceptor cells in the back of the retina. Activation
of rhodopsin instigates a cascade of biochemical reactions, which
eventually results in reduction of the steady (dark) current across
the photoreceptor plasma membrane. This is the photoreceptor response,
the signal that propagates to the brain enabling vision.
Employing an existing model for the biochemical cascade and the
response, expressed as a system of ordinary differential equations
involving 16 parameters, we present an approach based on statistical
sensitivity/uncertainty analysis and optimization,
to find parameters that produce a response matching experimental data.
AMS (MOS) Subject Classification. 92C45, 90C31, 62J02.
Phototransduction is the process by which light is converted into an
electrical response, in rod and cone photoreceptors in the retina.
A model for the cascade of biochemical reactions, and the ensuing
photoreceptor response, in rod photoreceptors of vertebrates has
been developed by (Hamer et al., 2003).
The cascade is described by 66 reactions involving 16 primary
parameters. The reactions can be translated into a system of
nonlinear ordinary differential equations (ODEs),
with coefficients involving the parameters (reaction rates).
Our goal is to find parameter values that produce a response
matching experimental data. This is a difficult, inverse
(hence ill-posed) problem that can be viewed as a multi-objective
optimization problem.
We employ a combination of statistical and optimization approaches
and tools to treat this parameter estimation problem.
To reduce the large 16-parameter search space, we use statistical
sensitivity analysis to identify the 4 most influential parameters
over which to optimize. To find "promising" starting values, we
use statistical sampling methods (Random and Lp-Tau, often employed
for uncertainty quantification). Evaluation of the cost function
requires execution of the forward model simulating the phototransduction
process; thus we must employ an efficient, derivative-free, nonlinear
optimizer. It turns out that the cost function is extremely "bumpy"
with multiple local minima. Nevertheless, we manage to identify
(several) parameter sets that produce responses reasonably close
to experimental data.
In §2 we briefly describe the phototransduction process,
the cascade of biochemical reactions, and the photoreceptor response.
In §3 we raise the issues arising in parameter optimization,
which are addressed in §4. We conclude with a summary in
§5.
Vision begins at photoreceptor cells in the back of the retina.
Photons are captured by rhodopsin molecules located on "discs"
(bilipid membranes) in the outer segment of (rod) photoreceptors.
Activation of rhodopsin instigates a cascade of biochemical reactions
(described below), the end product of which is production of PDE
(activated phosphodiestarase). PDE
hydrolyzes cGMP
(cyclic guanosime monophosphate), which diffuses in the cytosol
surrounding the discs, reducing its concentration.
The decrease of cGMP causes closure of some of the cGMP-gated channels
on the plasma membrane of the photoreceptor, resulting in lowering
the influx of positive ions, in particular Ca
, thus lowering
the local current
across the plasma membrane.
In darkness, the channels are open allowing influx of ions and a
steady dark current
is maintained by the
Na
/K
/Ca
exchanger. The reduction in current,
, known as the response, is the signal that
propagates to the brain enabling us to see.
Thus, the phototransduction process consists of two stages.
The first stage is the cascade of biochemical reactions,
leading from a photon captured by a rhodopsin molecule on a disc
at time
to production of activated PDE,
, at time
.
The entire cascade takes place on the activated disc.
The second stage takes place in the cytosol surrounding the discs.
Its input is
and, via diffusion of cGMP and Ca
in the
cytosol, results in drop of ionic current across the plasma membrane,
measured by the
The most detailed model for the cascade of reactions was developed
by (Hamer et al., 2003). It incorporates multi-stage shutoff of activated rhodopsin and
consists of 66 biochemical reactions, shown below.
Without explaining the meaning of each term, the cascade is activated
by rhodopsin (
), and the final product of interest is the
compount
, denoted simply
as PDE in the rest of the paper. Of the 66 reactions, only 50 actually
participate in the production of PDE
. They contain 16 primary
parameters (reaction rates), which we want to determine.
The reactions can be simulated in two different ways.
Directly, with a chemical kinetics stochastic simulator,
such as Dizzy (Dizzy, 2006; Ramsey et al., 2005)
or BioNetS (Adalsteinsson et al., 2004),
which perform Monte Carlo (Gillespie and other) simulations.
Or indirectly, by translating the reactions into ordinary differential
equations (ODEs) via mass action kinetics
(Patton, 2004), and solving the system of 50 nonlinear ODEs. This can be done very fast
in Fortran, which is particularly desirable for parameter estimation
as the code needs to be executed thousands of times.
We have verified that the average (of about 100) stochastic simulations
(which can be found conveniently with Dizzy)
agrees with the solution obtained from the system of ODEs.
The second stage of phototransduction, activated by
and
producing the photoreceptor response
, has been modeled at
various spatial resolutions: As 0-dimensional (bulk, well-stirred)
process by, among others,
(Pugh & Lamb, 2000; Hamer et al., 2003),
as 1-dimensional (longitudinal) process by
(Gray-Keller et al., 1999),
as 2-dimensional (axisymmetric) process by
(Khanal et al., 2004; Caruso et al., 2005; Alexiades & Khanal, 2007),
and
as 3-dimensional process (with incisures) by (Caruso et al., 2006).
Such models are discussed and compared in (Khanal & Alexiades, 2008)
in this volume.
Here we employ the bulk (0-dimensional) model of (Hamer et al., 2003),
which adds only four additional ODEs to the cascade model, and
allows us to compute the response
resulting from any set of
specified cascade parameters. In the sequel, we will refer to the code
solving the 50 ODEs for the cascade reactions and the 4 ODEs for
the response as the cascade code.
The 16 reaction rates (parameters) appearing in the cascade reactions
cannot be determined directly, or it would be enormously difficult
to do so experimentally as it would require measuring minute quantities
of participating species for each of the reactions. The only practical
way is to solve the inverse (ill-posed) problem of parameter estimation
by fitting a model to experimental data.
This involves an iterative optimization procedure of finding parameter
values that minimize an
norm of the difference between model
prediction and experimental data.
In phototransduction, the only reliably measurable quantity is the
response
of an isolated photoreceptor to light stimulation.
Recently, the technique has advanced to the point that response to
single-photon stimulation can be recorded.
Such Single Photon Response (SPR) experiments have been carried out on
salamander rod photoreceptors by Fred Rieke (Caruso et al., 2005).
Our goal is to determine cascade parameters that will produce a
response matching the experimental response data
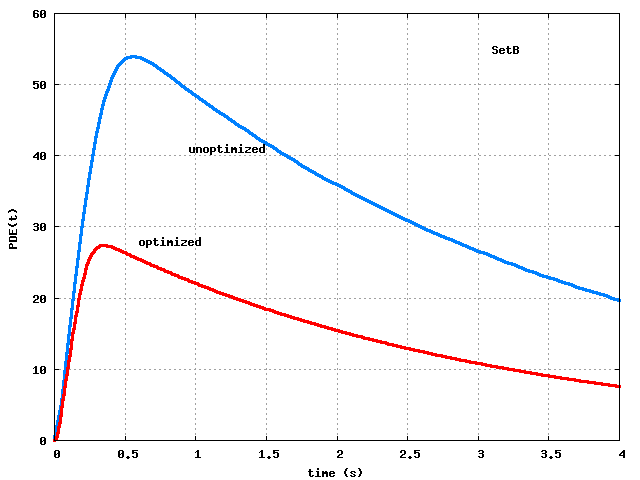
(wiggly solid curve in Fig.2
or Fig.3 or Fig.4).
Crucial features are the peak response (about 0.8%) and the time
at which the peak occurs (about 0.8 sec). For this, we estimate that
the cascade should produce a
curve with peak
at time
sec. Thus we want to find parameters
to drive the
and
,
produced by the cascade model, towards the target values 27
and 0.4 sec.
To this end, we set the target values
and
and seek parameters
to
The weights
allow unequal weighing of the two terms.
In such a problem, we are facing certain issues that need to be addressed.
Issue 1.Choice of optimizer:
Our objective function
in Eq.2 is highly nonlinear
and can be evaluated only by running the cascade code.
There is no explicit formula, and no derivatives are available for it.
Thus we need an efficient, nonlinear, derivative-free, global
optimizer. Powell's NEWUOA routine (Powell, 2004), written in Fortran,
meets these requirements and is simple to adapt and use.
Issue 2.Good starting values:
Hamer et al., 2003 published values for the parameters, which could be used as base values.
However, these produce
and
very far away from
our desired target values, and the optimizer cannot improve them much.
As in every nonlinear iteration, it is crucial to start with good
starting values. We employ statistical sampling
(SimLab 2.2; Saltelli et al., 2004),
to find promising starting parameters, as described in §4.
Issue 3.Large parameter space:
Optimizing over 16 parameters involves too large of a search space.
This calls for a parameter sensitivity study to figure out a
minimal number of most influential parameters and then optimize only over
those.
To address Issues 2. and 3. mentioned above, we employed
SimLab (SimLab 2.2; Saltelli et al., 2004),
an excellent and highly recommended simulation tool
for Uncertainty and Sensitivity Analysis, developed by italian
researchers for the European Commission.
4.1
Step 1. Good starting values via statistical sampling
SimLab can generate statistical samples by various statistical methods
(Ramdom, Latin Hypercube, QuasiRandom LpTau, Morris, Sobol).
For us, a 'sample' is a set of 16 parameter values.
For each of the 16 parameters, we specified a uniform distribution
over a range (appropriate for each parameter, see next subsection),
and generated 1000 samples by the Random sampling method,
1000 by the Latin Hypercube method,
and 1000 by the Quasirandom LpTau method.
We ran the cascade code on each sample (parameter set), to compute
the resulting
and
,
and looked for those that are reasonably near the target values.
A few promising ones were found, which we can use as starting parameters
for optimization. For illustration here, we present results for two such
"promising" parameter sets, labeled "SetA" and "SetB".
SetA was one of the samples generated by Random sampling,
while SetB was one of the LpTau samples.
To reduce the number of parameters over which to optimize, we performed
extensive sensitivity studies with SimLab to figure out which of the
16 parameters are most influential on
and
.
This involves generating a large number of samples (as for Step 1 above),
running the model (cascade code) on each sample, producing a file
containing the computed 'output variables'
(
and
in our case) in a format that SimLab can
read, and reading the file into SimLab (as 'external model output').
Then SimLab produces sensitivity indices and rankings according to
various statistical sensitivity measures, including the following:
PEAR: Pearson product moment correlation coefficient
SPEA: Spearman coefficient (PEAR on ranks)
PCC : Partial Correlation Coefficient
PRCC: Partial Rank Correlation Coefficient
SRC : Standardized Regression Coefficient
SRRC: Standardized Rank Regression Coefficient
For each of the variables
and
,
SimLab returns a table listing the parameters and their influence
rank according to each of the sensitivity methods. The rankings
are rather ambiguous. The methods do not necessarily assign the
same rank to a parameter, but they do agree on which parameters rank
first and second. The rankings depend strongly on the choice of
distribution for the parameters (uniform vs Gaussian, etc),
as well as on the choice of sampling method. Moreover, at least 1000
samples are required for reasonable agreement among methods.
Since we have no a priori knowledge about parameter distributions,
we assumed uniform distribution for each parameter over a wide range
about the values given by (Hamer et al., 2003). To address the uncertainty due to choice of sampling method,
we tried (1000 samples from) each of the three main sampling methods,
namely Random, Latin Hypercube, and LpTau, as mentioned above.
After many tries, we picked 4 of the 16 parameters that seemed to
influence
and
the most, which we shall refer to
as parameters
.
Only these four are varied during subsequent optimizations,
the rest being held fixed at their "promising" SetA or SetB values.
First, we optimize the quantity
in Eq.(2)
over the 4 influential parameters
,
trying to drive
and
towards the target values
and
.
We tried various combinations of weights and
,
or
seem to do best.
We observed high degree of non-uniqueness and ambiguity, and the
values that minimize
were usually not the best.
This is a typical problem in multi-objective optimization.
So we ended up using the optimizer as landscape explorer, by
printing out all the iterates to find those with best combination
of
and
. This can be done by
inspection, thanks to the efficiency of Powell's NEWUOA optimizer,
which explores the trust region within fewer than 30 iterations.
The best
we could find for SetA and SetB are plotted
in Fig.1, together with the
generated
by the initial (unoptimized) parameter values.
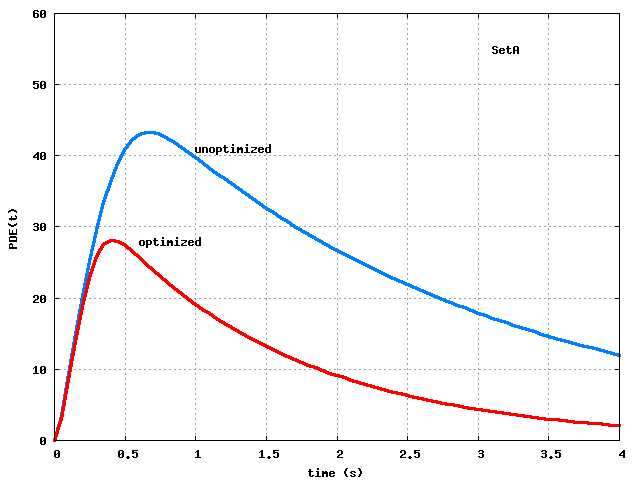
Clearly, the optimization process has greatly improved the peak,
and both sets approximate well the target values
,
.
Figure 1: Step 3 optimization of SetA and SetB parameters:
curves.
Both sets approximate well the targeted peak
(
,
).
However, when we look at the resulting response curves,
Fig.2,
the peak-optimized parameters produce lousy overall response
against the experimental data, even though they do well at the peak.
Figure 2: Step 3 optimization of SetA and SetB parameters:
curves.
Both sets do well at the peak but poorly afterwards.
Clearly, the strategy of optimizing to match only the peak is not sufficient.
4.4
Step 4. Optimization over the entire response history:
The failure of the peak-only optimization makes it necessary
to optimize over the entire history of the data !
That's bad news, as there are too many data points
(400, one every 10 msec), and the data curve
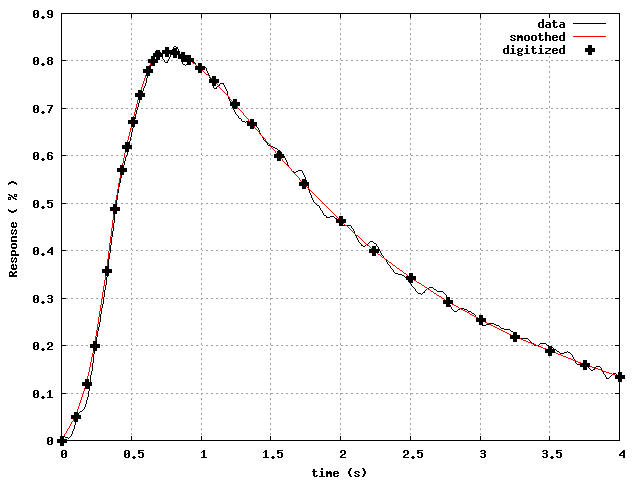
(solid curve in Fig.2) is too wiggly.
So, we first smoothed out the wiggly experimental data, and digitized
the smoothed curve using only 32 time points, as shown in
Fig.3.
Figure 3: Experimental data (wiggly curve), smoothed (dotted curve), and
digitized using only 32 points.
These digitized points were used as points
to minimize the L
-norm of
.
In fact, we seek (influential) parameters
that mininize the multi-objective cost function
with target values
,
now set to those of the
experimental response (0.82% and 0.7 sec, respectively), and
various weights
,
.
Note that the response values
,
,
as well as peak value,
, and the time it occurs,
,
are functions of the parameters
, in particular of
over which we are optimizing, and they can only
be found by running the cascade code.
The NEWUOA optimization code is efficient and typically uses
fewer than 100 evaluations of the cost function
, for
tolerance set to 2 %.
The multi-objective cost function
, natural as it is
for the problem, is far from ideal, unfortunately.
It turns out that many
can produce about the same value for
,
and the starting
is crucial, a nightmare for optimization.
Apparently, the landscape of
is extremely "bumpy",
with multiple, shallow, local minima.
Moreover, it is very hard to drive
lower (desirable),
and the last two terms in
seem to antagonize
each other; parameters that would lower
also
drive
away from the target value.
Under the circumstances, the choice
,
for weights
seems to do better than other combinations, yet leaves a lot to be
desired.
As in the peak-only case of Step 3, there is high degree of
non-uniqueness, and the values that minimize
were usually not the best.
So, again, we used the optimizer as landscape explorer, by
printing out all the iterates to find (by inspection) those with best
combination of
and
,
and then looking at the overall fit to the data.
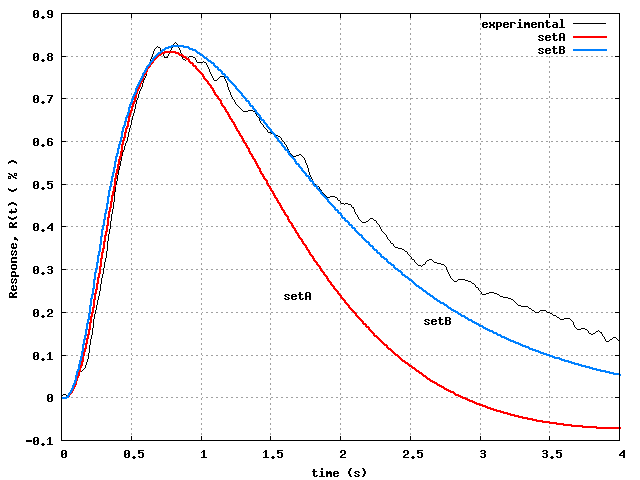
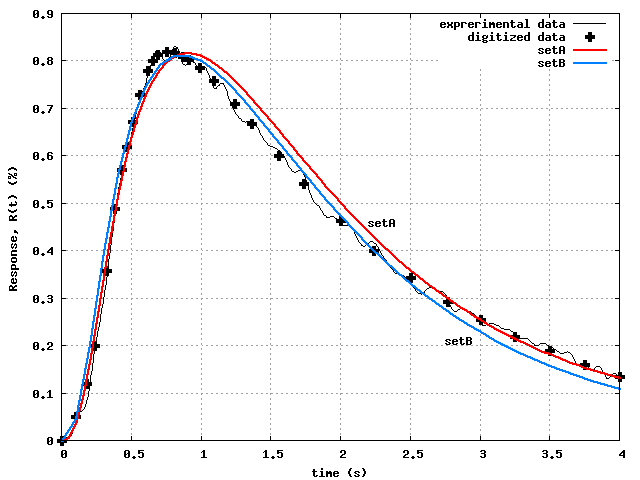
Fig.4 shows the best results we could obtain, after
many tries with various combinations of weights, tolerances,
and combinations of Step 3 and Step 4 optimizations. In fact,
the SetA curve was obtained by Step 4 optimization applied to the
Step 3 optimized SetA parameters, whereas the SetB curve was obtained
directly by Step 4 optimization of the unoptimized SetB parameters.
Considering the extreme natural variability of biological data,
the agreement of both SetA and SetB response curves with the
experimental data is very good.
Although both SetA and SetB do comparably well in capturing the
data, their optimized parameters are quite different, exhibiting the
severe non-uniqueness of the parameter estimation problem.
For example, the
optimized values of SetA are
347 , 618 , 0.258 , 0.051, while those of SetB are
190 , 741 , 0.279 , 0.108.
Viewed from another perspective, perhaps the values are not that
different.
A range of values for each parameter is perhaps the best one can
hope to determine on the basis of indirect biological data.
We discussed parameter estimation by fitting to experimental data
for phototransduction in rod photoreceptors of vertebrates.
This is a typical inverse (ill-posed) problem of parameter
identification that can be viewed as a multi-objective optimization
problem. The difficulties encountered and procedures to overcome
them were presented.
The difficulties include: large parameter space, highly nonlinear
dependence of quantities of interest on the sought parameters,
lack of good starting values, and high degree of non-uniqueness.
We employed statistical uncertainty and sensitivity analysis
(using the excellent SimLab software package),
to scope out the parameter space,
to reduce the number of parameters to only the most influential ones,
and to find "promising" starting parameter values for the subsequent
multi-objective optimization problem.
The ambiguities and pitfalls were pointed out.
The fact that quantities of interest can only be found via
simulation (running the forward model) necessitates using
an efficient, derivative-free optimizer. Powell's NEWUOA fulfills
these requirements.
Although natural for the problem, the multi-objective cost function
is far from ideal. Its landscape seems to be
extremely "bumpy", with multiple, shallow, local minima.
Thus we resorted to using the optimizer as landscape explorer, by
printing out the iterates to find (by inspection) those with best
combination of
and
,
and then looking at the overall fit to the data.
Despite the severe non-uniqueness and other difficulties encountered,
the approach turned out to be successful and we did find parameter
sets producing responses in good agreement with the available
experimental data. By biological standards, the agreement seen in
Fig.4 could be considered 'excellent'.
G Caruso, H Khanal, V Alexiades, F Rieke, HE Hamm, and E DiBenedetto,
Mathematical and Numerical Modeling of SpatioTemporal Signaling
in Rod Phototransduction,
IEE Proc. Systems Biology, 152(3) (2005) 119137.
G Caruso, P Bisegna, L Shen, D Andreucci, HE Hamm, and E DiBenedetto,
The Role of Incisures in Phototransduction in Vertebrate Photoreception,
Biophysical J., 91 (2006) 1192-1212.
Dizzy: chemical kinetics stochastic simulator in Java,
version 1.11.3 2006/04/18,
Institute for Systems Biology, Seattle WA,
http://magnet.systemsbiology.net/software/Dizzy/
M Gray-Keller, W Denk, B Shraiman, and PB Detwiler,
Longitudinal spread of second messenger signals in isolated rod
outer segments of lizards,
J. Physiology 519(3) (1999) 679-692.
RD Hamer, SC Nicholas, D Trachina, PA Liebman, and TD Lamb,
Multiple steps of phosphorylation of activated rhodopsin can
account for the reproducibility of vertebrate rod single-photon response,
J. Gen. Physiology 122 (2003) 419-444.
H Khanal, V Alexiades, and E DiBenedetto,
Response of Dark-adapted Retinal Rod Photoreceptors,
pp.138-145 in Dynamic Systems and Applications 4,
editor M Sambandham, Dynamic Publishers, 2004.
H Khanal and V Alexiades,
Models of Phototransduction in Rod Photoreceptors,
in Dynamic Systems and Applications 5,
editor M Sambandham, Dynamic Publishers, 2008 (this volume).
EN Pugh Jr. and TD Lamb,
Phototransduction in Vertebrate Rods and Cones,
Chapter 5, in DG Stavenga, WJ de Grip and EN Pugh Jr. (eds),
Handbook of Biological Physics, Vol. 3, pp.183-254. Elsevier 2000.
S Ramsey, D Orrell, and H Bolouri,
Dizzy: stochastic simulation of large-scale genetic regulatory networks,
J Bioinform Comput Biol. 3(2) (2005) 415-436.